
Dalam SparkNote pada Siklus Asam Sitrat, juga disebut Siklus Krebs, kami akan menjemput di mana kita tinggalkan di bagian terakhir dengan produk glikolisis aerobik, piruvat. Ketika oksigen hadir, piruvat bergerak keluar dari sitosol yang glikolisis berlangsung dan melintasi membran ke matriks mitokondria. Di sana, sebelum memasuki siklus asam sitrat yang tepat, piruvat mengalami tahap transisi, di mana dua pyruvates diubah menjadi asetil-koenzim dua A (asetil-KoA), dua molekul karbon dioksida, dan dua NADH. Kemudian, selama rangkaian delapan reaksi yang membentuk siklus asam sitrat, dua molekul asetil-KoA dioksidasi, menghasilkan dua molekul karbon dioksida lebih dan 2 ATP. Karbon dioksida yang dihasilkan dalam kedua proses adalah karbon dioksida kita keluarkan ketika kita bernapas.
Siklus asam sitrat, atau siklus Krebs, merupakan pusat metabolisme, karena pada tahap ini sebagian besar karbohidrat, lipid, dan protein terdegradasi oleh oksidasi. Salah satu ciri yang menandai siklus asam sitrat adalah bahwa hal itu tidak hanya memiliki fungsi degradatif. Sejumlah koenzim yang sangat penting diproduksi dalam reaksi siklus itu. Koenzim ini pergi untuk fosforilasi oksidatif, mengakibatkan hasil besar 32 ATP. Aspek lain yang menarik dari siklus asam sitrat adalah status sebagai "siklus": yang productof akhir siklus, oksaloasetat, adalah molekul yang diperlukan untuk reaksi pertama dari siklus dengan asetil-CoA.
Kita akan mulai pembahasan kita dengan melihat konversi piruvat ke asetil-KoA, bahan awal dari siklus asam sitrat. Berikutnya , kita akan mengikuti delapan reaksi siklus asam sitrat yang pada akhirnya mengarah pada produksi oksaloasetat dan koenzim berbagai yang pergi untuk digunakan dalam fosforilasi oksidatif.
Ketentuan
Asetil Koenzim A - Sebuah molekul kecil yang membawa kelompok-kelompok fungsional asetil dalam sel. Terdiri dari kelompok asetil melekat pada molekul A koenzim. Produk awal dari siklus asam sitrat.
Acontinase - Siklus asam sitrat Enzim yang bertanggung jawab untuk mengkatalisis reaksi yang mengubah asam sitrat untuk isocitrate dengan pelepasan air.
Adenisine trifosfat (ATP) - Molekul dari mana sel-sel memperoleh energi. Terdiri dari sebuah molekul adenisine terikat dengan tiga fosfat, masing-masing mengandung ikatan fosfat energi, terutama ikatan ketiga. Dengan melanggar ikatan satu dan mengurangi ATP untuk adenisine fosfat di (ADP), sel dapat mendapatkan energi untuk melaksanakan berbagai proses.
Alpha-ketoglutarat dehidrogenase - Siklus asam sitrat Enzim yang bertanggung jawab untuk mengkatalisis reaksi yang mengubah alfa-ketoglutarat untuk suksinil-KoA dengan pelepasan karbon dioksida dan produksi NADH dari NAD.
Amphibolic - Suatu istilah yang digunakan untuk menggambarkan suatu proses yang memiliki kedua sifat anabolik dan katabolik.
Anabolic - Jangka menggambarkan reaksi dikatalisis enzim-dalam sel yang melibatkan sintesis molekul kompleks dari subunit sederhana dan yang menggunakan energi.
Katabolik - Jangka menggambarkan reaksi dikatalisis enzim-dalam sel yang melibatkan degradasi molekul ke dalam subunit lebih sederhana dengan melepaskan energi.
Sitrat sintase - Siklus asam sitrat Enzim yang bertanggung jawab untuk mengkatalisis reaksi yang mengembun asetil-KoA dengan oksaloasetat untuk menghasilkan asam sitrat.
Siklus asam sitrat - Juga dikenal sebagai Siklus Krebs; jalur metabolik ditemukan pada organisme aerobik yang mengoksidasi gugus asetil KoA menjadi karbon dioksida dan air, menghasilkan 1 ATP, dan sejumlah koenzim yang memainkan peran penting dalam langkah berikutnya dari respirasi, fosforilasi oksidatif.
Koenzim A - Sebuah molekul yang digunakan untuk mentransfer enzimatis kelompok asetil dalam sel.
Dihydrolipoyl transasetilase - Satu dari tiga enzim yang membentuk kompleks piruvat dehidrogenase multienzim. Bertanggung jawab untuk mengkatalisis penambahan molekul KoA untuk struktur asetil-lipoamide.
Flavin adenin dinukleotida - Sebuah koenzim yang berpartisipasi dalam reaksi oksidasi dan reduksi dengan menerima dan menyumbangkan atom hidrogen. Terlibat khususnya dalam oksidasi dan pengurangan karbon-karbon ikatan ganda dan tripel.
Fumarase - Siklus asam sitrat Enzim yang bertanggung jawab untuk mengkatalisis reaksi yang mengubah fumarat L-malat dengan pelepasan air.
Glikolisis - Sebuah jalur metabolisme yang terjadi dalam sitosol sel. Glikolisis terdiri dari serangkaian reaksi yang diambil secara keseluruhan mengubah glukosa untuk mensintesis ATP dan piruvat.
Guanosin trifosfat - Sebuah molekul yang mirip baik dalam bentuk dan fungsi menjadi ATP.
Isocitrate dehidrogenase - Siklus asam sitrat Enzim yang bertanggung jawab untuk mengkatalisis reaksi yang mengubah isocitrate ke alfa-ketoglutarat dalam mekanisme dua-langkah yang mengarah pada produksi molekul NADH dan karbon dioksida.
Malat dehidrogenase - Siklus asam sitrat Enzim yang bertanggung jawab untuk mengkatalisis reaksi yang mengubah L-malat ke oksaloasetat dengan produksi molekul NADH.
Metabolisme - Semua reaksi yang terjadi dalam organisme yang berpartisipasi dalam akuisisi atau konversi energi untuk digunakan dalam organisme.
Oksaloasetat - Empat-molekul karbon yang mengikat dengan asetil KoA pada langkah pertama dari siklus asam sitrat untuk membentuk enam-karbon asam sitrat. Selama serangkaian reaksi, asam sitrat dipecah kembali menjadi dua 2 molekul CO dan oksaloasetat, memungkinkan untuk siklus untuk menjadi lagi sambil menghasilkan ATP dan sejumlah koenzim.
Oksidasi - Reaksi yang melibatkan hilangnya keseluruhan elektron dari molekul atau atom tertentu. Dapat terjadi dengan penambahan oksigen atau dengan penghapusan hidrogen.
Fosforilasi oksidatif - Sebuah proses yang terjadi dalam mitokondria yang menghasilkan pembentukan ATP dari aliran elektron ke oksigen.
Piruvat dehidrogenase - Satu dari tiga enzim yang membentuk kompleks piruvat dehidrogenase multienzim. Bertanggung jawab untuk menambahkan TPP untuk piruvat dengan pelepasan karbon dioksida.
Piruvat dehidrogenase kompleks multienzim - Sebuah nama menyeluruh diberikan kepada tiga enzim yang mengkatalisis reaksi yang mengubah piruvat ke asetil-KoA.
Pengurangan - Reaksi yang menghasilkan keuntungan keseluruhan elektron ke molekul tertentu atau atom. Dapat terjadi dengan penambahan atom hidrogen atau oleh penghapusan sebuah atom oksigen.
Suksinat dehidrogenase - Siklus asam sitrat Enzim yang bertanggung jawab untuk mengkatalisis reaksi yang mengubah suksinat untuk fumarat dengan produksi sebuah molekul FADH.
Suksinil-CoA sintetase - Siklus asam sitrat Enzim yang bertanggung jawab untuk mengkatalisis reaksi yang mengubah suksinil-KoA untuk suksinat.
Sebelum Siklus Asam Sitrat
Setelah muncul dari glikolisis, piruvat dua diangkut ke dalam mitokondria. Ada, piruvat menjalani tahap transisi sebelum memasuki siklus asam sitrat yang sebenarnya. Dalam fase ini piruvat diubah menjadi asetil koenzim-A produk (asetil-KoA), yang dimulai dalam siklus asam sitrat. ;
2 Piruvat + 2 koenzim A + 2NAD + -> 2 asetil-KoA 2 CO 2 + 2 NADH
Catatan: Siswa mengambil ujian biologi AP tidak perlu tahu lebih banyak tentang proses transisi ini.
Pembentukan Asetil-CoA
Asetil-CoA adalah produk umum dari karbohidrat, lipid, dan pemecahan protein. Ini terdiri dari kelompok asetil melekat pada molekul A koenzim. Koenzim A adalah molekul besar yang berisi molekul ADP dengan kelompok dua sisi rantai yang berasal dari lengan fosfat. Kelompok asetil melekat pada ujung rantai ini samping. Dengan cara ini, koenzim A bertindak sebagai pembawa gugus asetil. Ketika dipecah oleh air, sejumlah besar energi dilepaskan, yang, seperti akan kita lihat, drive siklus asam sitrat. Cara yang paling umum yang asetil-KoA yang diturunkan dalam jalur metabolik adalah dengan bantuan dari kompleks piruvat dehidrogenase multienzim.
Para multienzim piruvat dehidrogenase adalah kompleks dari tiga enzim yang berbeda yang bersama-sama mengubah piruvat menjadi asetil-KoA dengan bantuan sebuah molekul koenzim A dan NAD. Mekanisme untuk pembentukan asetil-KoA yang kompleks, seperti terlihat di bawah ini. Umumnya, dalam reaksi 1, piruvat dehidrogenase enzim menarik sebuah molekul karbon dioksida dari piruvat. Hal ini dilakukan dengan bantuan molekul yang disebut TPP yang membentuk ikatan sementara dengan molekul piruvat. Karbon dioksida reaksi penghapusan mirip dengan yang dari dekarboksilase piruvat ragi dalam fermentasi alkohol.

Dalam reaksi 2, transasetilase dihydrolipoyl enzim membantu untuk melampirkan molekul lain sementara disebut suatu lipoamide. Dengan pembentukan ikatan, molekul TPP dari langkah pertama dilepaskan mengarah pada pembentukan sebuah kelompok asetil. Pada langkah ketiga, kelompok ini lipoamide berkurang dan dirilis sebagai molekul KoA serangan kelompok asetil. Kami sekarang memiliki asetil-KoA. Enzim ketiga, dehidrogenase dihydrolipoyl, bertanggung jawab untuk memulihkan lipoamide ke aslinya, negara yang teroksidasi sehingga dapat digunakan kembali dalam siklus dalam langkah keempat. Molekul NAD menegaskan dirinya pada titik ini, membantu reoxidize lipoamide tersebut.
Pada titik ini, kami telah asetil-KoA dan siap untuk memasuki siklus asam sitrat.
Reaksi Siklus Asam Sitrat
Kami sekarang siap untuk mulai pergi melalui reaksi-reaksi siklus asam sitrat. Siklus dimulai dengan reaksi antara asetil-KoA dan oksaloasetat karbon empat untuk membentuk enam-karbon asam sitrat. Melalui langkah-langkah berikutnya dari siklus, dua dari enam karbon asam sitrat meninggalkan sebagai karbon dioksida untuk akhirnya menghasilkan produk karbon empat, oksaloasetat, yang digunakan kembali pada langkah pertama dari siklus berikutnya. Selama delapan reaksi yang terjadi, untuk setiap molekul asetil-KoA siklus memproduksi tiga NADH dan satu dinukleotida flavin adenin (FAD/FADH2), bersama dengan satu molekul ATP.
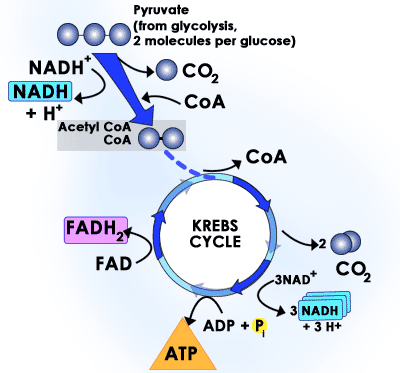
Catatan: Siswa mengambil tes AP umumnya tidak perlu lebih lanjut tentang spesifik dari siklus asam sitrat dari apa yang terkandung dalam gambar di atas dan ayat.
Reaksi 1: Synthase Sitrat
Reaksi pertama dari siklus asam sitrat dikatalisis oleh enzim sitrat sintase. Dalam langkah ini, oksaloasetat ini bergabung dengan asetil-KoA untuk membentuk asam sitrat. Setelah dua molekul bergabung, sebuah molekul air menyerang asetil mengarah ke rilis koenzim A dari kompleks.

Reaksi 2: Acontinase
Reaksi berikutnya dari siklus asam sitrat dikatalisis oleh enzim acontinase. Dalam reaksi ini, molekul air akan dihapus dari asam sitrat dan kemudian menempatkan kembali di lokasi lain. Efek keseluruhan dari konversi ini adalah bahwa gugus-OH dipindahkan dari 3 'ke 4' posisi pada molekul. Transformasi ini menghasilkan isocitrate molekul.

Reaksi 3: dehidrogenase Isocitrate
Dua peristiwa terjadi dalam reaksi 3 dari siklus asam sitrat. Dalam reaksi pertama, kita melihat generasi pertama kami NADH dari NAD. Enzim dehidrogenase isocitrate mengkatalisis oksidasi dari gugus-OH pada posisi 4 'dari isocitrate untuk menghasilkan perantara yang kemudian memiliki molekul karbon dioksida dihapus dari itu untuk menghasilkan alpha-ketoglutarat.

Reaksi 4: Alpha-ketoglutarat deydrogenase
Dalam reaksi 4 dari siklus asam sitrat, alpha-ketoglutarat kehilangan sebuah molekul karbon dioksida dan koenzim A ditambahkan pada tempatnya. Dekarboksilasi terjadi dengan bantuan NAD, yang diubah menjadi NADH. Enzim yang mengkatalisis reaksi ini adalah alpha-ketoglutarat dehidrogenase. Mekanisme konversi ini sangat mirip dengan apa yang terjadi dalam beberapa langkah pertama dari metabolisme piruvat . Molekul yang dihasilkan disebut suksinil-KoA.

Reaksi 5: suksinil-CoA sintetase
Para sintetase suksinil-KoA enzim mengkatalisis reaksi kelima dari siklus asam sitrat. Pada langkah ini molekul guanosin trifosfat (GTP) disintesis. GTP adalah molekul yang sangat mirip dalam struktur dan sifat energik menjadi ATP dan dapat digunakan dalam sel-sel dalam banyak cara yang sama. GTP sintesis terjadi dengan penambahan gugus fosfat bebas untuk molekul PDB (mirip dengan sintesis ATP dari ADP). Dalam reaksi ini, gugus fosfat bebas pertama serangan molekul suksinil-KoA melepaskan KoA. Setelah fosfat melekat pada molekul, itu dipindahkan ke PDB untuk membentuk GTP. Produk yang dihasilkan adalah suksinat molekul.

Reaksi 6: dehidrogenase suksinat
Suksinat dehidrogenase enzim mengkatalisis pemindahan hidrogen dari dua suksinat dalam reaksi keenam dari siklus asam sitrat. Dalam reaksi, sebuah molekul FAD, koenzim mirip dengan NAD, direduksi menjadi FADH2 karena mengambil hidrogen dari suksinat. Produk dari reaksi ini adalah fumarat.

FAD, seperti NAD, adalah bentuk teroksidasi sementara FADH2 adalah bentuk tereduksi. Meskipun FAD dan NAD melakukan peran oksidatif dan reduktif yang sama dalam reaksi, FAD dan bekerja NAD pada kelas yang berbeda dari molekul. FAD mengoksidasi karbon-karbon ikatan ganda dan triple, sementara sebagian besar NAD mengoksidasi karbon-oksigen obligasi.
Reaksi 7: fumarase
Dalam reaksi ini, enzim mengkatalisis fumarase penambahan molekul air untuk fumarat dalam bentuk sebuah gugus-OH untuk menghasilkan molekul L-malat.

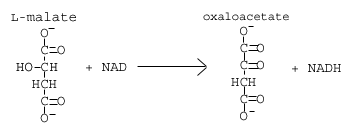
Reaksi 8: Malate dehidrogenase
Dalam reaksi akhir dari siklus asam sitrat, kita regenerasi oksaloasetat dengan mengoksidasi L-malat dengan molekul NAD untuk menghasilkan NADH.
Kesimpulan
Kami sekarang telah menyimpulkan diskusi kita tentang reaksi yang membentuk siklus asam sitrat. Hal ini membantu pada saat ini untuk mengambil satu menit untuk mengambil stok apa siklus asam sitrat telah dihasilkan dari satu molekul asetil-KoA.
The-asetil KoA, telah dioksidasi menjadi dua molekul karbon dioksida.
Tiga molekul NAD dikurangi menjadi NADH.
Satu molekul FAD dikurangi menjadi FADH2.
Satu molekul GTP (setara dengan ATP) diproduksi.
Perlu diingat bahwa pengurangan adalah benar-benar mendapat elektron. Dengan kata lain, NADH dan FADH2 molekul bertindak sebagai pembawa elektron dan digunakan untuk menghasilkan ATP dalam tahap berikutnya dari metabolisme glukosa, fosforilasi oksidatif. Dalam SparkNote berikutnya pada Fosforilasi oksidatif dan rantai transpor elektron , kita akan belajar apa proses berlangsung untuk akhirnya memperoleh mayoritas dari ATP yang kita butuhkan untuk bahan bakar aktivitas kita sehari-hari.





















